熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
漿果攝氧量
使用克拉克型氧氣微呼吸電極OX-MR和微呼吸系統(Unisense)測量漿果的氧氣吸收量。每個重復由九個漿果組成。在裝有9個分離漿果的測量室中注入通氣的Milli-Q水,不斷攪拌并在25 C的水浴中保持恒溫。測量室內水中的[O2]至少持續15分鐘,每隔5秒讀取一次數據,根據[O2]的下降斜率確定穩定的呼吸速率。
漿果乙醇濃度
用液態N2冷卻的A11基本研磨機將每個重復的10個冷凍漿果研磨成細粉。按照生產商的說明使用乙醇檢測試劑盒對乙醇進行定量。簡而言之,乙醇脫氫酶(ADH)催化乙醇氧化成乙醛。然后,乙醛在醛脫氫酶(AL-DH)和NAD+的作用下進一步氧化成乙酸和NADH。用FLUOstar Omega平板閱讀器在340納米波長下測量NADH的形成。
使用比重計測量漿果孔隙度
從第二季所有處理的每個重復中抽取三個新鮮漿果作為子樣品。將小心剪去花梗的漿果浸沒在脫氣的Milli-Q水中,然后使用真空泵和干燥器在-740 mm Hg下抽真空。每抽真空270秒,干燥器降壓30秒。總作用時間為50分鐘。在抽真空前后在抽真空之前和之后獲得。使用Hubbard-Carmick比重瓶(25 mL)估算單個漿果的體積。漿果內空氣空間的體積是假定真空滲透50分鐘后,所有空氣空間都充滿水而估算的。50分鐘后,在真空條件下無法從漿果中觀察到明顯的氣泡流。通過計算內部空氣空間體積(v2)從而計算出漿果的孔隙率。
X射線顯微CT斷層掃描確定完整漿果的內部空氣空間體積
為了確認使用比重計測量的漿果孔隙度,并對漿果的內部氣隙結構進行成像,使用了X射線顯微CT來估計漿果成熟過程中漿果內部氣隙體積和分布的變化。文中使用Skyscan 1076顯微鏡對第二季中A+NI和A+I處理的每個重復的一個漿果進行了成像。采用59 kV、149μA、Al 0.5 mm濾光片、2356 ms曝光、0.4旋轉步長采集整個漿果(果梗附著)的二維投影。使用NRecon(bruker-microct.com)獲得漿果的橫向灰度圖像,圖像像素大小為8.5μm(相當于15μm空間分辨率或3×10-6 mm3象素大小)。結果表明,這種分辨率能夠充分評估蘋果的多孔微觀結構。為了對A+I處理的漿果中的空氣空間進行三維可視化,使用CT分析儀(bruker-microct.com)對體積進行了大津閾值化處理,并使用去斑功能只接受超過500或1000體素的連續體積作為連接的空氣空間。使用CTvox(bruker-microct.com)將僅包含氣隙的反轉圖像重建為三維漿果圖像。在計算I型和NI型葡萄樹漿果的子房體積時,參照灰度圖像進行手動閾值處理,并使用CT分析儀進行去斑設置,只接受超過1000體素的連續體積作為子房體積。使用CT分析儀計算腦室體積。
結果
生長條件和物候
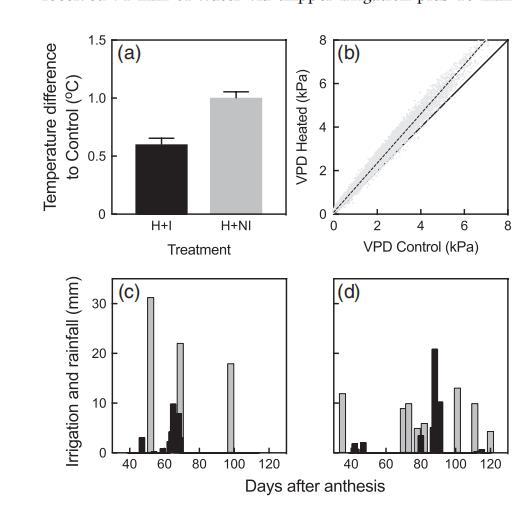
圖1a、第二季(S2)中,升溫灌溉(H+I)和升溫非灌溉(H+NI)與常溫灌溉(A+I)葡萄樹花后40天(DAA)至收獲期的樹冠環境溫度差異。
圖1b、升溫(H+I)與對照(A+I)葡萄樹冠層蒸氣壓差的比較.
圖1c(S1)和d(S2)從40 DAA到采收期間的日降雨量(黑色)和灌溉量(灰色)。
圖1c,d總結了水分投入情況。在第1季,2014年12月至2015年2月期間,I型葡萄樹通過灌溉獲得了71毫米的水量,外加10毫米的有效降雨量,而NI型葡萄樹則獲得了10毫米的有效降雨量(圖1c)。在70 DAA附近記錄到頻繁的降雨事件(圖1c)。在第二季中,2015年12月至2016年2月期間,I型葡萄樹通過灌溉獲得了74毫米的灌溉水量和31毫米的有效降雨量(圖1d)。在87至91 DAA期間出現了一系列大規模降雨(圖1d)。
在第2季的40至125 DAA期間,常溫灌溉處理的日最高環境溫度為21.2至44.5攝氏度,而日最低溫度為6.3至22.8攝氏度。同期,升溫灌溉的日最高氣溫平均升高0.6攝氏度,升溫非灌溉的日最高氣溫平均升高1.0攝氏度(圖1a),但對最低氣溫沒有影響。灌溉型葡萄樹較大的樹冠削弱了被動升溫的效果。VPD的升高與最高溫度的升高成正比(圖1b)。
莖干水勢、氣孔導度和氣體交換
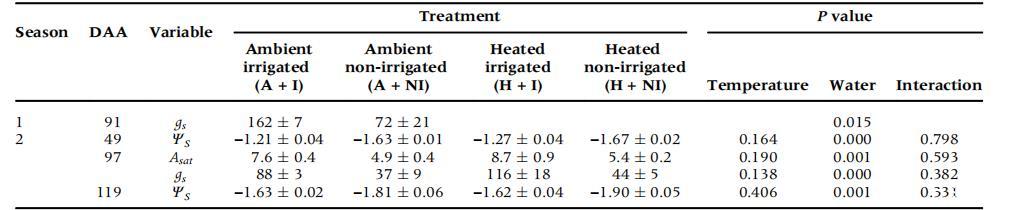
表1、兩個季節處理下的水分脅迫生理指標和方差分析結果。
缺乏灌溉會導致明顯的水分脅迫生理指標(表1)。在第一季,在第91 DAA天的晴朗天氣測量了氣孔導度[平均光合有效輻射(PAR)=2852±98μmol/(m2*s)],此時非灌溉葡萄藤的gs是灌溉葡萄藤的一半,接近作為水分脅迫指標的50 mmol/(m2*s)臨界值。非灌溉葡萄藤測得相對較低的gs與該地點以前在開花后測得的結果相似。
在第二季,中午的莖干水勢(ψs)反映了對NI葡萄藤的水分脅迫。升溫葡萄藤沒有表現出水分脅迫,在氣體交換性狀或ψs方面與灌溉沒有相互作用(表1)。在第二季中,有兩次觀察到NI藤蔓的gs受到類似的抑制,與A+I藤蔓相比,光飽和時的二氧化碳凈同化(Asat)顯著降低(表1)。
漿果成熟
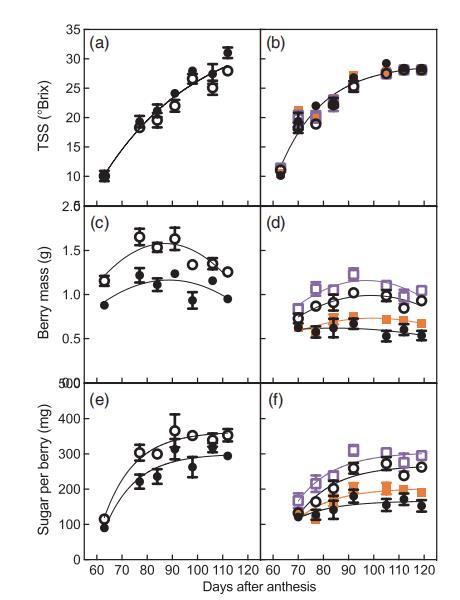
圖2、第1季漿果成熟期(a,c,e)和第2季漿果成熟期(b,d,f)顯示(a,b)總懸浮固體含量、(c,d)漿果鮮重和(e,f)每粒漿果含糖量。處理為:常溫灌溉(A+I)(白圈)、常溫非灌溉(A+NI)(黑圈)、升溫和灌溉(H+I)(白方塊)、升溫和非灌溉(H+NI)(橙方塊)。若顯示不同的擬合線,則表明擬合線之間存在顯著差異。
圖2比較了第1季和第2季不同處理的漿果總懸浮固體含量、漿果鮮重和每顆漿果含糖量的動態變化。第1季中A+I和A+NI葡萄樹的漿果質量峰值分別在在87 DAA和89 DAA;在第2季,質量峰值出現在99 DAA(A+I)、83 DAA(A+NI)、97 DAA(H+I)和100 DAA(H+NI)(圖2c、d)。對于每個漿果的含糖量,不同處理下的糖積累速率常數沒有明顯差異(圖2e、f)。
漿果活體組織
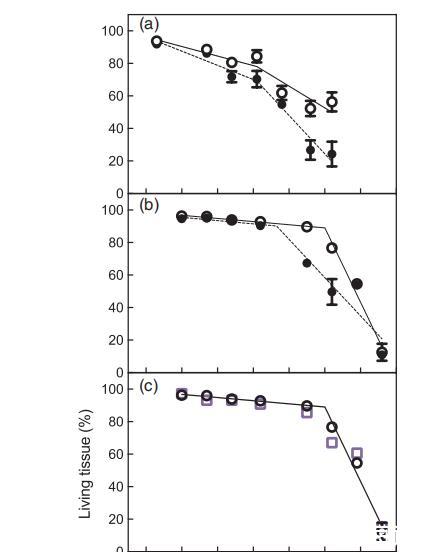
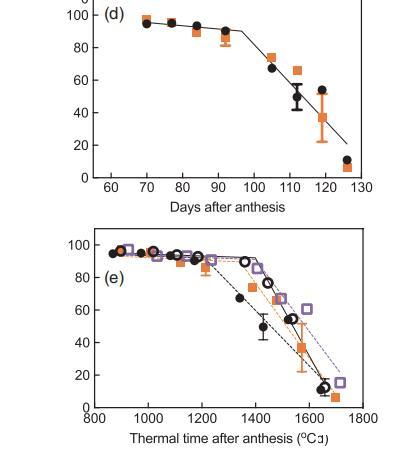
圖3、第1季(a)和第2季(b-d)從花期開始的一段時間內漿果活體組織的比例,以及從花期開始的熱量時間內(e)比較以下處理:常溫灌溉(A+I)(白圈)、常溫非灌溉(A+NI)(黑圈)、升溫灌溉(H+I)(白方塊)、升溫非灌溉(H+NI)(橙方塊)。為清楚起見,(c)中重復了A+I,以便與H+I進行比較;(d)中重復了A+NI,以便與H+NI進行比較。
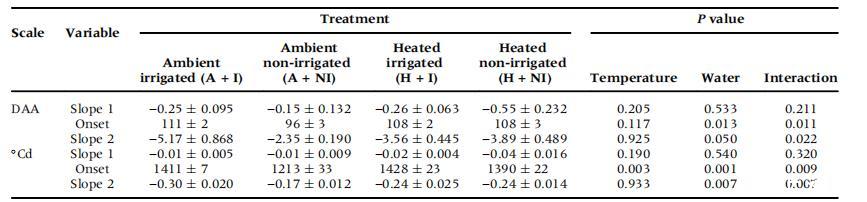
表2、設拉子第2季節CD快速開始前(斜率1)和之后(斜率2)的細胞死亡率,以及方差分析的結果。
圖3顯示了兩季中不同處理對CD動態變化的影響。分段線性回歸確定了發育閾值之后的慢CD階段(斜率1)和快CD階段(斜率2)(Bonada等人,2013b)。第1季中,A+I和A+NI漿果的快速CD階段開始時間相同,均為90-91 DAA,但在第2季中,A+NI葡萄樹的快速CD階段開始時間較早,但CD率較低(表2)。快速成熟閾值的提前出現與漿果質量的峰值相對應。按時間順序考察,升溫對快速成熟期的開始或發展速度沒有影響(圖3c,d),但這兩個參數與灌溉有顯著的交互作用(表2)。
將第2季的CD與花期開始的熱時間作圖(圖3e),結果表明,與A+I葡萄藤相比,A+NI葡萄藤的活體組織開始快速衰退的時間更早,快速階段(斜率2)也更小(表2)。升溫增加了CD快速階段開始的熱時間,并且與灌溉有顯著的交互作用(表2)。缺水減緩了未升溫葡萄植株活體組織的快速衰退(斜率2),且與溫度有顯著的交互作用(表2)。在快速階段開始之前,處理或交互作用對CD緩慢階段的斜率沒有影響(表2)。
 相關新聞
相關新聞